![]()
Conferences / Workshops ( 2000~2011 ) / Seminars and Group Meetings
2010 NCTS September Workshop on Critical Phenomena and Complex Systems
|
|
|
|
|
|
|||||||
|
|
Date : |
10-11 September 2010 |
|
|
|||||||
|
|
Place : |
The auditorium on 1st floor, Institute of Physics, Academia Sinica, Taipei |
|
||||||||
| Organized by : |
National Center for Theoretical Sciences (Critical Phenomena and Complex Systems focus group) Institute of Physics, Academia Sinica (Taipei)
|
||||||||||
| Contact Info. : |
Miss Chia-Chi Liu (Secretary,
Physics Division, NCTS)
|
||||||||||
|
|
Speakers : |
||||||||||
|
Mr. Norifumi Fujikane Graduate School of Science, and Cybermedia Center, Osaka University, JAPAN E-mail: |
|||||||||||
|
(Poster) The Transmission Characteristics of OV Model We study a complex dynamical behavior of linearly-coupled interacting elements expressed by a variation of the optimal velocity model (OV model). OV model [1] was originally proposed as a model of traffic flow. It provides us with a unified description of both the free flow and the congested flow by a single set of dynamical equations. The model is based on an idea that a vehicle accelerates or decelerates to the optimal velocity which depends on the headway distance to its preceding vehicle. The function which determines the velocity is called the optimal velocity function (OV function). Since the model is based on a second order differential equation, inertia induces effective delay in adjustment to the optimal velocity. In this work, we consider OV model as a prototype model of interacting elements, not necessarily of vehicle traffic. Each element takes a single variable x, and an element follows its preceding element through OV model:
where the parameter a is the sensitivity and ∆xi is difference in x of i-th and i-1-th elements. We employ OV function f(∆x) of the following form:
The parameter
α
is the maximum value of the optimal velocity. In contrast to the
traffic flow model, negative values are allowed for ∆xi in
this model. We arrange the elements in a linear chain and analyze
transmission characteristics of a periodic motion in case that the first
element is subjected to a periodic driving with amplitude A and
frequency ω.
References: [1] M. Bando, K. Hasebe, A. Nakayama, A. Shibata and Y. Sugiyama, Dynamical model of traffic congestion and numerical simulation, Phys. Rev. E 51, 1035 (1995).
|
|||||||||||
|
|||||||||||
|
The Glassy State of Biopolymers: Why It's Important? An important aspect of the protein physics is the low temperature glass transition experimentally observed at approximately 200 K [1-8]. The understanding of this transition is achieved by combining information obtained via rather different methods: micro-mechanical experiments [1], NMR [2], Moessbauer spectroscopy [3], calorimetric studies [4], pressure release experiments [5], and X-ray scattering of synchrotron radiation [6]; see [7, 8] for reviews. Although there still open important questions here, the rough physical picture of the glass transition in globular proteins is constructed in close analogy to glass forming liquids [7, 8] and synthetic polymers [9, 10]. In particular, it is believed that the large-scale conformational motion of proteins freezes at (approximately) 200 K, analogously to freezing of cooperative motion in glass-forming liquids [7, 8] and segmental motion in synthetic polymers [9, 10].Thus, the glassy features as such are not important in the native state of globular proteins at physiological temperatures, though the glass transition at much lower temperatures can be still employed for gaining some understanding on the relevant motion in proteins [7, 8].Here we shall demonstrate via micro-mechanical methods that the native type I collagen fibril (made of fibrous protein, type I collagen triple-helices) is in a glassy state at physiological temperatures. This state is displayed via frequency-dependent visco-elastic characteristics (the Young's modulus and the decrement of damping) of the native fibril. Upon heating the fibril goes out of the glassy state at temperatures around 70 C, a phenomenon known as the softening transition [9, 10] (in contrast to the proper glass transition, where the system enters into the glassy state upon cooling). Approximately in the same temperature interval the fibril starts to undergo the denaturation process [13-18]; this fact might explain why the glassy state of the native fibril went so far unnoticed in the vast literature devoted to collagen. We stress that the glassy state is a feature of the native fibril, and it is absent for the heat-denatured fibril, as seen below.
References: [1] V.N. Morozov and S.G. Gevorkian, Biopolymers 24, 1785 (1985). [2] A. G. Palmer III, J. Williams and A. McDermott, Phys.Chem. 100, 13293 (1996). [3] E. W. Knapp, S. F. Fisher and F. Parak, J. Chem. Phys. 78, 4701 (1983). [4] G. Sartor, E. Mayer and G. P. Johari, Biophys. J. 66, 249 (1994). [5] I.E.T. Iben et al., Phys. Rev. Lett. 62, 1916 (1989). [6] K. Achterhold et al., Eur. Biophys. J. 25, 43 (1996); C. Keppler et al., Eur. Biophys. J. 25, 221 (1997). [7] H. Frauenfelder, S. G. Sligar, P. G.Wolynes, Science 254, 1598 (1991). [8] D. Ringe and G. A. Petsko, Biophysical Chemistry 105,667 (2003). [9] J. D. Ferry, Viscoelastic Properties of Polymers (John Wiley & Sons, New York, 1980). [10] G. M. Bartenev and S. Ya. Frenkel, Physics of Polymers (Khimia, Leningrad, 1990) (In Russian). [11] Collagen, Structure and Mechanics, ed. by P. Fratzl (Springer, Berlin, 2008). [12] J. Engel and H. P. Bachinger, Top. Curr. Chem. 247, 7 (2005) [13] A. R. Haly and J. W. Snaith, Biopolymers 10, 1681 (1971). A. Finch and D. A. Ledward, BBA 278, 43 (1972). [14] C. A. Miles, T. V. Burjanadze and A. J. Bailey, J. Mol. Biol. 245, 437 (1995). [15] E. I. Tiktopulo and A. V. Kajava, Biochemistry 37, 8147 (1998). [16] C. A. Miles et al., Biophysical Journal 76, 3243 (1999). C. A. Miles and T. V. Burjanadze, Biophysical Journal 80, 1480 (2001). [17] G. I. Tsereteli et al., Journal of Thermal Analysis and Calorimetry 62, 89 (2000). [18] S. G. Gevorkian et al., Phys. Rev. Lett. 102, 048101 (2009).
|
|||||||||||
|
|||||||||||
|
Thermodynamics and Kinetics
of the Long Charged Macromolecule in the Entropic Trap
|
|||||||||||
|
|||||||||||
|
Finding Disease Modules in Protein Interaction Networks As molecular networks represent the backbone of molecular activity within cells, integrative analysis of transcriptomic profiles in the context of protein interaction networks provides opportunities for understanding the molecular mechanism of diseases. While protein-protein interaction data constitute static network maps, integration of condition-specific co-expression information provides clues to the dynamic features of the networks. Here, we present a network-based comparative analysis that integrates gene expression profiles with protein-protein interaction and biological function annotations to elucidate heart failure related molecular modules. The revealed hub genes and molecular modules may be used as potential disease markers and provide new directions for heart failure therapy. |
|||||||||||
|
|||||||||||
|
Asymptotic Expansion for the Resistance between Two Maximally Separated Nodes on a Finite Resistor Network
We analyze the
exact formulae for the resistance between two arbitrary notes in a
rectangular network of resistors under free, periodic and cylindrical
boundary conditions obtained by Wu [J. Phys. A 37, 6653 (2004)]. Based
on such expression, we then derive the exact asymptotic expansions of
the resistance between two maximum separated nodes on an M x N
rectangular network of resistors with resistors r and s in the two
spatial directions. The all coefficients in this expansion are expressed
through analytical functions. |
|||||||||||
|
|||||||||||
|
Application of REM in Strings, Conformal Models and Financial Markets: Exact Results We mapped exactly off critical string models into Random Energy Model (REM) by Derrida and deduced some critical dimensions. We mapped any 2-d conformal model into REM like model. The scaling of 2-point and 3-point correlation functions are exactly calculated in REM approach.
Evolution Models in Multi-Dimensional Spaces We consider a fitness landscape where the fitness depends on the mean number of alleletypes at several fractions of genome. We calculate the mean fitness for the general fitness case. The recombination model is formulated for such a fitness landscape.
|
|||||||||||
|
|||||||||||
|
(Poster) A Lattice Model of Intrinsically Disordered Proteins Intrinsically disordered proteins (IDPs) are proteins characterized by lack of stable three-dimensional structures in their free states[1]. Under the existence of their targets, however, IDPs fold into unique three-dimensional structures upon binding to the targets. This process is called coupled folding and binding. IDPs have many functions like transcriptional regulation and signal transduction by the process. We have constructed a simple lattice model of IDPs to simulate coupled folding and binding and to discuss the advantages of coupled folding and binding. Our model includes an IDP and its target in a finite system. Because of the simplicity of the model and a powerful Monte Carlo method [2,3], we can calculate free-energy landscapes at an arbitrary temperature. We discuss a mechanism of coupled folding and binding based on the free-energy landscape and a role of the flexibility of IDPs on their biological function.
References: [1] H. J. Dyson and P. E. Wright. Nature Rev. Mol. Cell Biol. 6, 197 (2005). [2] Y. Iba, G. Chikenji, and M. Kikuchi. J. Phys. Soc. Jan 67, 3327 (1998). [3] G. Chikenji, M. Kikuchi, and Y. Iba. Phys. Rev. Lett. 83, 1886 (1999).
|
|||||||||||
|
Mr. Hiroshi Sugahara Graduate School of Frontier Biosciences, and Cybermedia Center, Osaka University, JAPAN E-mail: |
|||||||||||
|
(Poster) Relaxing Neutrality of a Neutral model on a Square Lattice Hubbell's neutral model of biodiversity[1] has been criticized for its neutrality assumption that all individuals belonging to different species have the same birth, death and dispersal rates. It predicts species abundance distributions and several empirical tests support it. However, why neutral model provides a good approximation although it is likely that ecological communities are often not neutral? Further work is needed to assess in which situations the neutral model is a good approximation. Recently, Zhou et al. showed that the results of a nearly neutral model with even slight difference of birth rates are very different from those of the neutral model[2]. We investigated a nearly neutral model on a square lattice by changing not only birth rates but death rates. The square lattice model enables us to treat dispersal limitation more realistically. In our model, the probabilities of death and birth rates of species i are given by
where di and bi are the death and birth rates at the individual level and ni is the abundance of species i, respectively, and the sum of j is taken over all communities and that of k is over a dispersal kernel.
We distributed
two species with different death and birth rates on a square lattice and
calculated the median of the time to fixation(MTF).
References: [1] S. P. Hubbell, The Unified Neutral Theory of Biodiversity and Biogeography. Princeton, NJ: Princeton University Press (2001). [2] Shu-Rong Zhou, Da-Yong Zhang, A nearly neutral model of biodiversity, Ecology 89(1), 248, (2008).
|
|||||||||||
|
Prof. Kei Tokita Cybermedia Center, Graduate School of Science, and Graduate School of Frontier Biosciences, Osaka University, JAPAN E-mail: tokita@cmc.osaka-u.ac.jp |
|||||||||||
|
Random Matrices and Their Application to Mathematical Biology Matrices with elements drawn randomly from statistical distributions are called random matrices. Since a successful application of the random matrix to nuclear physics, it has been established that physical properties of many disordered systems (such as amorphous materials, a magnetic alloy like spin glasses, chaos, etc.) are determined by mathematical properties of random matrices, in particular, by their eigenvectors and eigenvalues [1-3]. One of the most famous results is the so-called Wigner's semicircular law [1] which states that the average density ρ(x) of the eigenvalue x of a NXN symmetric real random matrix aij in the limit of matrix size N→∞ is
where each element aij is drawn from independent identical distribution with zero odd-order moments, finite even-order moments and variance σ2. The above theorem (the circular version for asymmetric real random matrix) was applied to mathematical biology at comparatively early days and one of the most significant results was given by May [4]. He considered a large-dimensional ecological equation with various types of interspecies interactions, e.g. prey-predator relations, competition, mutualism, parasitism, etc. The interspecies interactions were represented by an N-dimensional random matrix {aij} whose diagonal elements are -1 and whose off-diagonal elements are assigned as Gaussian random real numbers with mean 0 and variance α2 (with probability C, the connectance) or 0 (with probability 1-C). He found a kind of phase transition from stability to instability in which the equilibrium solution corresponding to the coexistence of all species becomes unstable if α>(NC)-1/2, the phenomenon of which is first discovered numerically by Gardner and Ashby [5]. This result clearly exhibits that an ecological system with complex interspecies interactions is not stable if any of the system size (N), the connectance (C) or the variance (α2) of the random matrix is large enough, in other words, if the system is "complex", which challenged a proposition that the larger and the more complex communities are more stable than simple ones, widely supported by field ecologists by 1960s. After May's work and the clear conclusions he reached, a number of mathematical biologists attempted to explain the discrepancy between the observed complexity of ecosystems in nature and the results of these mathematical studies. Community structure in which the author is interested is symmetry and correlation of elements of random matrix although only fully asymmetric random matrix was considered in May's analysis where the elements aij and aji are independent each other. Sommers et al [6] extended the Wigner's semicircular law to a general SXS real random matrix with correlations between off-diagonal elements as,
where the overline denotes random average over the distribution of the random matrix's elements as
if the elements obeys Gaussian distribution with mean 0 and variance 1. The parameter ν and γ are the variance and the level of symmetry respectively. For γ=1 the random matrix is fully symmetric, for γ=0 aij and aji are uncorrelated and for γ=-1 the matrix is fully antisymmetric (prey-predator relations in the term of ecology). The author also interested in the global behavior of a nonlinear dynamics of community ecology while only linear stability was studied in May's analysis. One of the most analyzed one is the replicator equation,
where xi(t) denotes population of the i-th species. I present recent results of the analysis on the replicator equations, in special, with random correlated interactions ({aij}) defined by Eqs. (1). Statistical mechanics of random systems enables us to treat the system analytically and phase diagram of stability and the species abundance distributions of the system with the parameters ν and γ are presented [7, 8].
Acknowledgements: The author thanks to the collaboration and the illuminating discussions with Tobias Galla (University of Manchester) and Yoshimi Yoshino (University of Tokyo). The present study is supported by grants-in-aid from MEXT, Japan (No. 14740232 and 17540383) and through the priority area `Systems Genomics'.
References:
[1] E. P. Wigner, On the
distribution of the roots of certain symmetric matrices, Ann. Math.
67, 325, (1958).
|
|||||||||||
[Academia Sinica] [Institute of Physics] [Library of the Institute of Physics] [Electronic Journals] [Related Links]
Laboratory of Statistical and Computational Physics
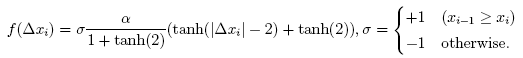
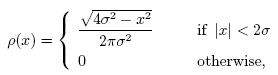
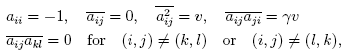 (1)
(1)